今年の初め頃、David Sherman と Kendal Houk らのグループから hapalindole 生合成での Cope 転移を触媒する酵素 HpiC1 の計算化学も含めた酵素解析の論文が発表されました。
この論文のメインは X 線結晶構造に基づいた酵素機能の解析になりますが、 Houk らの計算結果も論文中で非常にポイントとなっており面白いと思いましたので、簡単に紹介したいと思います。
“Structural basis of the Cope rearrangement and cyclization in hapalindole biogenesis”
Sean A. Newmister, Shasha Li, Marc Garcia-Borràs, Jacob N. Sanders, Song Yang, Andrew N. Lowell, Fengan Yu, Janet L. Smith, Robert M. Williams, K. N. Houk and David H. Sherman
Nat. Chem. Biol.2018,14, 345–351. DOI:10.1038/s41589-018-0003-x
概要
{kind=link}
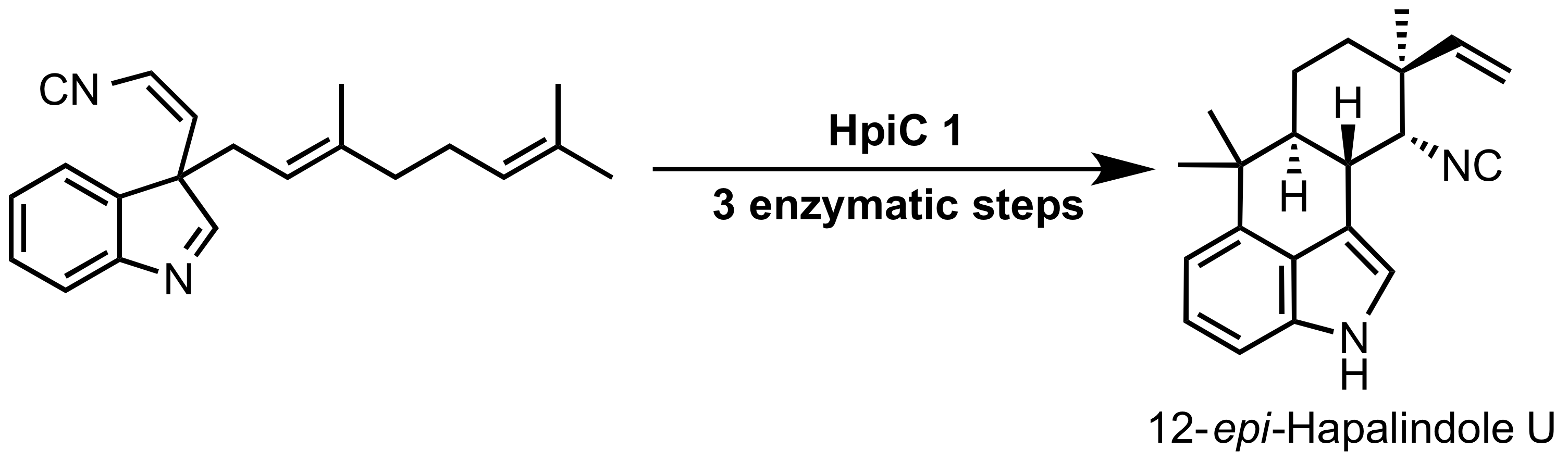
Hapalindole alkaloids は、多様な多環性構造および多様な生物学的活性が特徴的なシアノバクテリアが生産する非常に構造多様性に富んだ天然物群です。これらの複合代謝産物は、共通の生合成中間体から Stig cyclases によって次の 3 つのステップで合成されます:(i) 珍しい Cope 転位、(ii) 6-exo-trig 環化、および (iii) 芳香族求電子置換反応。 本論文は、in vitro での 12-epi-Hapalindole U の生成を触媒する Stig cyclase である HpiC1 の構造解析の報告です。1.5 Å の解像度で得られた結晶構造から、HpiC1 がモノマー当たり 2 つの
計算手法
基質のコンフォメーション探索には、Schrödinger MacroModel を使用し、最もエネルギーの低いものを用いて DFT 計算を行なったようです。やはり、計算の仕方が丁寧です。
DFT 計算には Gaussian09 A.02 が用いられました(何故 revision A なのかは不明。g16 A.02 の書き間違い?)。B3LYP/6-31G(d) で構造最適化が行われ、B3LYP/6-31++G(d,p), IEFPCM diethyl ether solvent (
Docking には Auto Dock Vina が使われています。exhaustiveness 以外は、全て default 設定とのことです。MD シミュレーションには、AMBER16 が使われています。
内容
Hapalindole alkaloids のようにインドールにプレニル基がついた天然物の生成は、prenyl transferase によって触媒されます。そして、この indole + prenyl の構造は、様々な転移反応や複雑な環化反応を起こしやすく、天然物の構造多様性を作り出す一因となっていると言っても過言ではありません(参考文献 4)。また、prenyl transferase も大変面白い酵素であり、有機合成的なインドールの反応性に関係なく、インドールの様々な位置をプレニル化することができ、また付加するプレニル基の数も決まっておりません。
自然界のマイルドな条件下で Cope 転移、と聞くと珍しいと思う人もいるかもしれませんが、これまでにも [1,5] シグマトロピー転位(参考文献 5)や Cope 転移(参考文献 6)などが天然物の生合成過程で進行することが報告されてきました。これまでにも類似の報告がありながら、なぜ今回の論文が NCB に掲載されたかというと、Stig cyclaseの初の結晶構造という点に尽きると思います。また、変異実験や MD 計算の結果が素晴らしく一致している点や も大きなポイントだと思います。
Asp214
著者らは、これまでに報告のある Stig cyclase の酵素の配列を比較し、全ての Stig cyclase で保存されている残基である Asp214 に注目しました。よく保存されているということは、酵素活性に必須である可能性が高いからです。
本論文の変異体酵素を用いた実験では、Asp214 を Ala に変えてしまうと全く反応が進行せず原料回収になってしまっているため、ますます触媒残基である可能性が高まってきました。
また、FimC5 という同様の基質から同様の反応を経て 12-epi-fischerindole を生産する酵素との配列比較で Ser 138 が重要そうだということも予想されました。Y101F/F138S という変異酵素の解析を行うと 12-epi-fischerindole が唯一の生成物として得られてくることが分かりました。つまり HpiC1 の F138 が生成物の切り替えに重要であることが示唆されました。
DFT 計算
{kind=link}
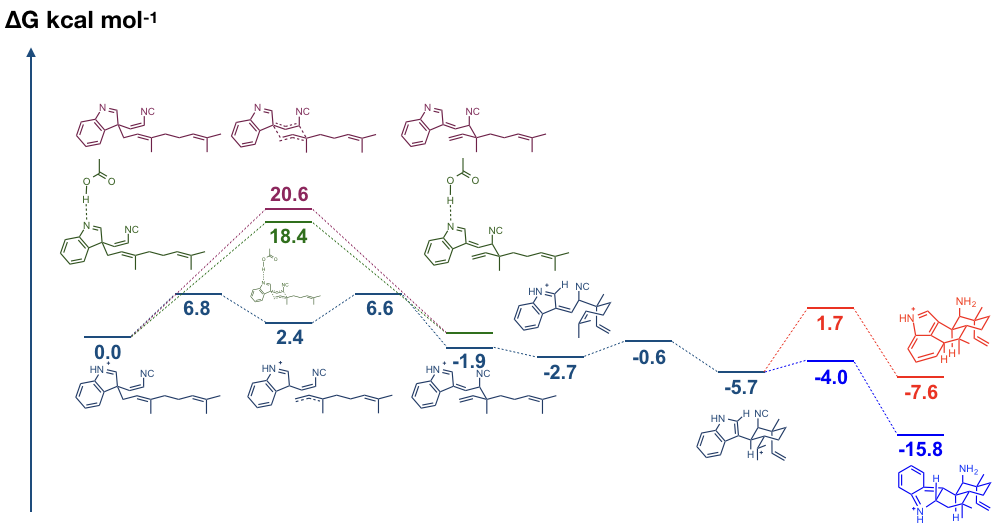
この Cope 転移ですが、理論的には 4 種類の遷移状態があります(二つの chair 型と 二つの boat 型)。しかし、そのうちの一つしか今回の生成物の立体化学と一致するものはありません。
これらの結果をより詳しく解析するために DFT 計算が行われました。中性条件での計算、indolenine の N をプロトン化したもの、Asp のモデルとして酢酸を含めた theozyme 計算の 3 通りが計算されています。
上図に示しますように Theozyme 計算の結果を見ると中性条件の計算に比べて約 2.2 kcal/mol エネルギー障壁が低くなっています。これにより反応速は約 40 倍早くなります。この結果は、Asn214 が Cope 転移を加速させていることを強く支持するものとなっています。
Cope 転移の後には、6-exo-trig cyclization が進行します。この反応は indolenine の N がプロトン化されている時のみ反応が進行するということが DFT 計算により示されました。プロトン化されていない状態だと、負に帯電した indolenine の C3 と正に帯電した C16 が反応して cyclobutane を形成してしまうようです。ここまでの計算結果より、プロトン化が非常に重要であることが示されており、Asp214 がこの反応に深く関わっているのではないかということを強く支持しています。
最後に、芳香族求核置換反応が進行します。二つの異なる遷移状態を経て、それぞれ別々の化合物が生成します。上図で赤色で示したものが 12-epi-Hapalindole U の前駆体であり、青色で示したものが 12-epi-Fischerindole U の前駆体です。
このくらいであれば、誰にでもできそうな DFT 計算ですが、Houk らはきちんと indolenine の N がどれくらい安定化されているかという点について触れています。ただエネルギーを算出するだけでなく、しっかりとした考察が素晴らしい点なのだと思います。
MD 計算
今回の記事ではあまり詳しく触れませんでしたが、本論文では MD 計算がかなり良い仕事をしています。各変異体酵素の変異させた残基が基質近傍にあるかどうか、活性部位の内側を向いているかなどを明らかにしたり、基質の R 体 と S 体のどちらの方が酵素に認識されやすいか?Y101F/F138S HpiC1 でなぜ生成物が変わるのか?なども明らかにしています。
参考文献
- “Structural basis of the Cope rearrangement and cyclization in hapalindole biogenesis”
Sean A. Newmister, Shasha Li, Marc Garcia-Borràs, Jacob N. Sanders, Song Yang, Andrew N. Lowell, Fengan Yu, Janet L. Smith, Robert M. Williams, K. N. Houk and David H. Sherman
Nat. Chem. Biol. 2018,14, 345–351. DOI:10.1038/s41589-018-0003-x - “On the dielectric “constant” of proteins: smooth dielectric for macromolecular modeling and its implementation in DelPhi.” Li, L., Li, C., Zhang, Z. & Alexov, E. J. Chem. Theory Comput. 2013, 9, 2126–2136.
- “What are the dielectric “constants” of proteins and how to validate electrostatic models?”
Schutz, C. N. & Warshel, A. Proteins 2001, 44, 400–417. - “Mechanistic studies on the indole prenyltransferases” Tanner, M. E. Nat. Prod. Rep. 2015, 32, 88–101.
- “Isochorismate pyruvate lyase: a pericyclic reaction mechanism?”
DeClue, M. S., Baldridge, K. K., Künzler, D. E., Kast, P. & Hilvert, D. J. Am. Chem. Soc. 2005, 127, 15002–15003. - “A Cope rearrangement in the reaction catalyzed by dimethylallyltryptophan synthase?”
Luk, L. Y., Qian, Q. & Tanner, M. E. J. Am. Chem. Soc. 2011, 133, 12342–12345.
関連する記事
- Coenzyme B12 依存性酵素 ONIOM 計算の汎関数ベンチマーク
- Spinosyn A の生合成 〜[4 + 2] or [6 + 4] Cycloaddition?〜
- 遷移状態後の枝分かれでの Dynamic Effect
- ダブルハイブリッド密度汎関数理論 doubly hybrid density al theory
- 正宗・Bergman 環化の計算化学【エンジイン】
- 【スパコン】Titan【アメリカ】
- gaussian scan の結果を解析するプログラム
- 原始地球上の非生物的条件下での核酸塩基の生成機構
- GRRM の振動計算の結果を Gauss View で解析する方法
- 【スパコン】天河二号【中国】
- GPU を用いた並列計算
- Rh(II) 触媒を用いたインドール合成の計算化学【反応機構解析】
- 構造最適化の閾値は、何を意味しているのか?
- 量子化学計算で a.u. を使う理由 〜Why Atomic Unit?〜